Pigtrop - Le centre d'information sur l'élevage porcin et les filières porcines des pays du Sud
;)
Processing methods of cassava leaves | The effects of simple processing methods of cassava leaves on HCN content and intake by growing pigs
The HCN content in leaves can be reduced by sun-drying and ensiling. The use of simpler methods of processing the fresh leaves, such as chopping, washing and / or wilting would facilitate the use of cassava leaves as animal feed (with the permission of Livestock Research for Rural Development).
Cassava (Manihot esculenta, Crant) is cultivated in Viet Nam in an area exceeding 250,000 ha (Bui van Chinh and Le Viet Ly 2001). Traditionally the crop is grown for the roots, which are used as human and animal food and industrially as a source of starch. In recent years, attention has been focused on the leaves as feed for ruminants (Preston 2001;Wanapat 2001) and pigs (Bui Huy Nhu Phuc et al 2001).
The main limiting factor to the use of cassava leaves as animal feed is the presence of cyanogenic glucosides, which give rise to hydrocyanic acid (HCN) when the plant tissue is broken down by processing or during ingestion by animals. The cyanide levels in leaves are influenced by genetic, physiological, edaphic and climatic differences with the stage of maturity being perhaps the major source of variation (Ravindran 1995).
Many researches have demonstrated that the HCN content in leaves can be reduced by sun-drying (Bui Van Chinh and Le Viet Ly 2001) and ensiling (Ly and Rodríguez 2001). However, sun-drying is difficult when cassava leaves are harvested in the wet season and ensiling is labour-intensive. The use of simpler methods of processing the fresh leaves, such as chopping, washing and / or wilting would facilitate the use of cassava leaves as animal feed.
Materials and Methods
Effect of age of leaves on chemical composition
Twenty varieties of cassava were grown under the same condition in the research garden of the Crop Production faculty in Hue university. The experimental design was a randomized block with plots of 2*5 m and three replications of each variety. Cassava stem cuttings were used as planting material at 50*30 cm spacing between rows and cuttings. Cattle manure was applied at 800 kg/ha prior to planting. Samples of leaves were taken at the time of root harvest (180 days) from the tender green leaves at the top of the stem. Determinations were made of dry matter (DM), crude protein and HCN according to AOAC (1990) procedures.
Effect of chopping and wilting on HCN content
Leaves from the high-yielding cassava variety (Cao san) were collected at 9.00 am. The petioles were removed, and the leaves chopped into small pieces (2-3 cm) and then spread out on a plastic sheet under a roof and allowed to wilt until 9.00 the next morning. Samples were taken for analysis immediately after collecting the leaves (09.00 am) and again at 12.00 am, 15.00 pm, 18.00 pm and at 09.00 am the following morning.
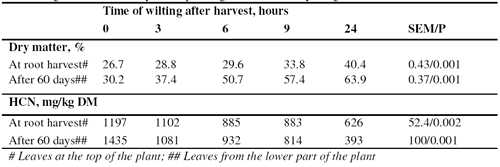
Samples of cassava leaves were taken after 60 days of growth and at the time of harvesting the roots after 180 days. For the 60-day growth (CL60), the leaves were collected at a point on the stem about 2/3 of the height of the plant. The leaves taken at the harvesting of the root (CL180) were from the top of the cassava tree.
Feed intake of cassava leaves by growing pigs
Treatments
The three treatments were:
The basal diet was a mixture of ensiled cassava root and rice bran (2:1 ratio in fresh basis) with an estimated crude protein content of 6.2% in DM.
Animals and experimental design
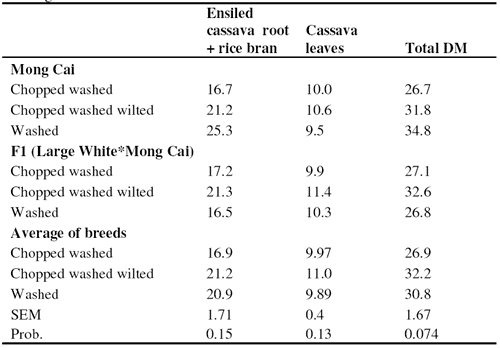
There were 3 pigs of each of two breeds (Mong Cai and F1 Large White*Mong Cai), with average initial weight of about 23 kg. They were allocated to the three treatments in an arrangement of two Latin squares (2[3*3]) with periods of 14 days (Table 1).

Feeds and feeding system
The cassava leaves without petiole were collected at the time of root harvesting (180 days). For the "W" treatment, they were washed two times in a plastic basin (each time for about 5 minutes with 10 litres water per 2 kg of leaves). For "CW" the leaves were chopped into small pieces (2-3 cm) and then washed as in "W". For "CWW" processing was as for "CW" followed by wilting over-night under a roof.
The ensiled cassava roots were made from whole cassava roots purchased from a local farmer. They were washed, ground and mixed with 0.5% NaCl prior to ensiling in plastic bags under anaerobic conditions for 21 days. Rice bran was purchased from the market. Prior to feeding, the ensiled cassava root and the rice bran were mixed in a ratio of 2:1 (fresh basis).
During a 7-day adaptation period, the cassava leaves and the mixture of ensiled cassava root and rice bran were fed ad libitum in separate feed troughs to establish the expected level of intake of the ensiled cassava root and rice bran mixture.
In the subsequent experimental period of 21 days the offer level of the mixture of ensiled cassava root / rice bran was set at 80% of the intake observed during the period of adaptation, while the cassava leaves continued to be fed ad libitum, both feeds in separate feed troughs. The mixture of ensiled cassava root / rice bran was fed three times daily at 06.00, 12.00 and 18.00 hr. The cassava leaves were given 4 times at 08.00, 10.00, 14.00 and 16.00 hr.
Measurements
Records were kept daily of amounts of feed offered and refused. Representative samples were taken for analysis of DM, N and HCN according to AOAC (1990) procedures.
Results and discussion
Chemical composition of twenty cassava varieties
The composition of the leaves of twenty cassava varieties, taken from the upper part of the plant at the time of root harvesting, ranged from 23.7 to 31.1% for DM, 23.7 to 29.5% for crude protein (in DM) and 610 to 1840 for HCN (mg/kg DM) (Table 2). The range for crude protein was less than that reported by Ravindran (1995) (16.7 to 39.9%) but similar to that described by Bui Huy Nhu Phuc et al (2001) for cassava varieties in the South of Viet Nam (21 to 34 %). Other researchers have also shown that there is considerable variation in the chemical composition among leaves of different cassava varieties (Ravindran 1990; Eggum 1970).
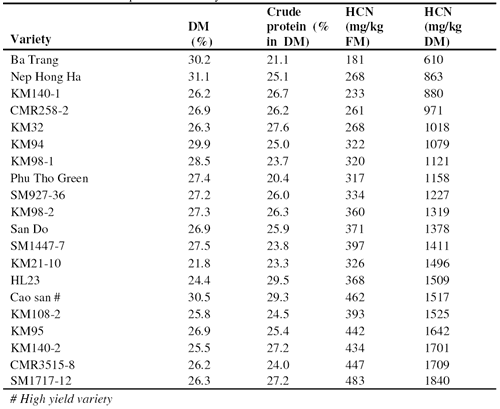
The protein in cassava leaves has an acceptable array of essential amino acids and compares favourably with alfalfa and soybean meal (Phuc and Lindberg 2001). However, the major concern with the use of cassava leaves as animal feed, is the risk of cyanide toxicity. In this study there was a wide variation in HCN content from 610 to 1840 mg/kg DM. The wide variation observed in the HCN content of cassava leaves has been attributed to genetic, physiological, edaphic and climatic differences (Gomez and Valdivieso 1985). In this study the environmental conditions (soil, fertilization and climate) were similar, thus it is probable that the major factor causing variation in HCN content was genetic. Pham Sy Tiep and Nguyen Van Dong (1998) reported that the HCN content of the leaves at the time of root harvest varied according to age and maturity: 442 (mg/kg fresh matter) in the green leaves at the top of the plant, 365 in green leaves below the top, 42.9 in mature leaves and 14.4 in the oldest leaves. In the present study the leaves taken for analysis were still very green, and the average HCN level (349 mg/kg fresh matter) was similar to that reported by Pham Sy Tiep and Nguyen Van Dong (1998). Duong Thanh Liem (1998) also reported similar values (from 305 to 425 mg/kg fresh leaves).
Effect of chopping and wilting on HCN content
The 58% reduction in the HCN content of the cassava leaves after wilting 24 hours in the shade (Table 3 and Figure 1) is similar to that (60%) reported by Bui Van Chinh and Le Viet Ly (1996), but much less than when wilting is under sunlight when up to 90% reduction in HCN has been reported (Gomez and Valdivieso 1985; Ravindran et al 1987). The faster rate of drying of the leaves sampled from the 60-day growth compared with the leaves from the mature plant (Figure 2) was not reflected in the changes in HCN content, which were similar for both sources of leaves.
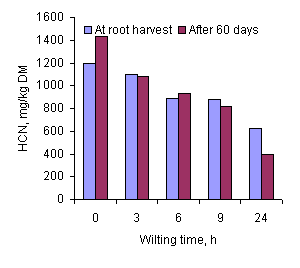
Feed intake of fresh cassava leaves by pigs.
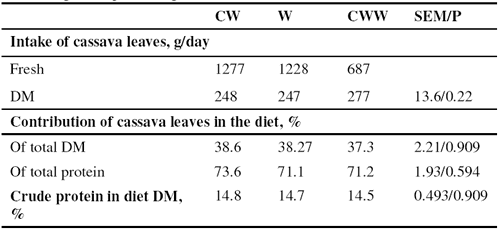
Washing alone or washing after chopping the leaves reduced slightly (by 16 and 21%) the HCN level (Table 4); however, the pink color in the urine observed when fresh cassava leaves were fed was no longer visible when the leaves were washed prior to feeding. Wilting for 24 hours after chopping and washing reduced the HCN by 82%.
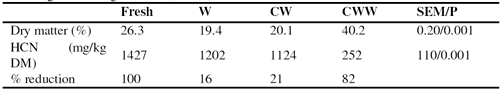
Although there were major differences in the HCN content of the cassava leaves (Table 3) as between washing with and without chopping and 24 hour wilting, this did not appear to affect the DM intake neither of the ensiled cassava root / rice bran mixture nor the cassava leaves (Table 5). The intake of cassava leaves accounted for over 30% of the total DM intake (Figure 2), resulting in intakes of HCN of 373 and 337 mg/day with washed and chopped / washed leaves, compared with 146mg/day for the wilted leaves (Table 6). Reported toxic levels of HCN (mg/kg live weight) for pigs are 1.4 (Getter and Baine 1938), 2.1 to 2.3 (Johnson and Ramond 1965), 4.4 (Butler 1973) and 3.5 (Tewe 1995). In the present study the HCN intakes were much higher (from 6.0 to 15 mg/kg live weight) yet no signs of toxicity were observed.
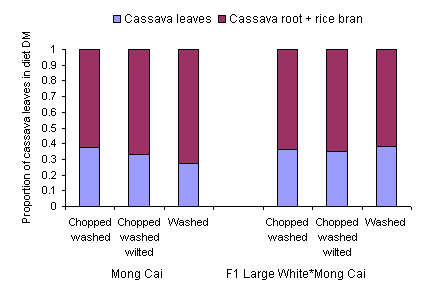
The cassava leaves contributed 37 to 39% of the diet DM and from 71 to 74% of the total protein, providing an overall level of 15% crude protein in the diet DM (Table 6). These levels of dietary crude protein provided by fresh cassava leaves are much higher than has previously been reported in the literature (Bui Huy Nhu Phuc 2001; Ly and Rodríguez 2001). However, they are comparable with results of a recent experiment in Cambodia (Chhay Ty and Preston 2005) in which fresh cassava leaves supplied 42% of the diet DM and 70% of the protein in a basal diet of broken rice fed to growing pigs.
Conclusions
* Pigs given restricted amounts (80% of observed voluntary intake) of a 2:1 mixture (fresh basis) of ensiled cassava roots and rice bran, and free access to fresh cassava leaves, which had been washed, chopped and washed or wilted for 24 hours, consumed similar amounts of total DM (range of 27 to 32 g DM/kg live weight.
* Levels of HCN were reduced slightly (16%) by washing and substantially (82%) by wilting, resulting in intakes of HCN from 6.0 to 15 mg/kg live weight, levels considerably higher than previously reported to be safe levels to avoid toxicity (1.4 to 4.4 mg/kg live weight).
* The fresh cassava leaves were readily consumed providing38% of the dietary DM and over 70% of the dietary protein.
References
- AOAC 1990 Official method of analysis, 13th edition Association of Official Analytical Chemists, Washington DC.
- Bui Huy Nhu Phuc, Ogle B and Lindberg J K 2001 Nutritive value of cassava leaves for monogastric animals; Proceedings of workshop on "Use of cassava as animal feed", Khon Kaen University, Thailand http://www.mekarn.org/procKK/phuc.htm
- Bui van Chinh and Le Viet Ly 2001 Study on the processing and use of cassava tops as animal feed. http://www.mekarn.org/prockk/chinh.htm
- ButlerG W 1973 Physiological and genetic aspects of cyanogenesis in cassava and other plants, Chronic cassava toxicity. Proceedings of the Interdisciplinary Workshop, London England, 29-30 Jan., 1973. IDRC -010e, pp. 65-71.
- Chhay Ty and Preston T R 2005: Effect of water spinach and fresh cassava leaves on intake, digestibility and N retention in growing pigs; Livestock Research for Rural Development. Vol. 17, Art. #14 Retrieved, from http://www.cipav.org.co/lrrd/lrrd17/2/chha17014.htm
- CIAT 1970 Swine nutrition unit annual report. CIAT, Cali, Colombia.
- Duong Thanh Liem 1998 Study on the use of different kinds of leaf meal for broiler chickens Agricultural Science and Technology, Hanoi pp: 165-167.
- Eggum O L 1970 The protein quality of cassava leaves. British Journal of Nutrition 24:761-769.
- Getter A O and Baine J 1938 Research on cyanide detoxification. Amer. J. Med. Sci., pp. 185-189.
- Gomez G and Valdivieso M 1985 Cassava foliage: chemical composition, cyanide content and effect of drying on cyanide elimination. Journal of the Science of Food and Agriculture 36: 433-441.
- Johnson R M and Ramond W D 1965 The chemical composition of some Tropical food plants. Manioc. Tropical Science 7, pp. 109-115.
- Ly J and Rodríguez L 2001 Studies on the nutritive value of ensiled cassava leaves for pigs in Cambodia. Proceedings of workshop on "Use of cassava as animal feed", Khon Kaen University, Thailand http://www.mekarn.org/prockk/ly.htm
- Phuc B H N and Lindberg J E 2001 Ileal digestibility of amino acids in growing pigs fed a cassava root meal diet with inclusion of cassava leaves, leucaena leaves and groundnut foliage. Anim. Sci. 72: 511-517.
- Preston T R 2001 Potential of cassava in integrated farming systems. Proceedings of workshop on "Use of cassava as animal feed", Khon Kaen University, Thailand http://www.mekarn.org/prockk/pres.htm
- Ravindran V 1990 Feeding value and digestibility of cassava leaf meal for growing pigs. Proceedings Fifth Australasian Animal Production Congress. Volume 3, Taipei, Taiwan, p.20.
- Ravindran V 1995 Preparation of cassava leaf products and their use as animal feeds. In: Roots, tubers, plantains and bananas in animal feeding, FAO Animal Production and Health Paper, No 95. pp. 11-116.
- Ravindran V, Kornegay E T and Rajaguru A S B 1987 Influence of processing methods and storage time on the cyanide potential of cassava leaf meal. Animal Feed Science and Technology 17: 227-234.
- Tewe O O 1995 Detoxification of cassava products and effects of residual toxins on consuming animals. In: Roots, tubers, plantains and bananas in animal feeding. FAO Animal Production and Health Paper No 95, pp. 81-95.
- Wanapat M 2001 Role of cassava hay as animal feed in the tropics. Proceedings of workshop on "Use of cassava as animal feed", Khon Kaen University, Thailand http://www.mekarn.org/prockk/wana3.htm
Citation of this paper
Commentaires
CIRAD © 2007 (Tous droits réservés) - Informations légales - Page mise à jour : 13/04/2007